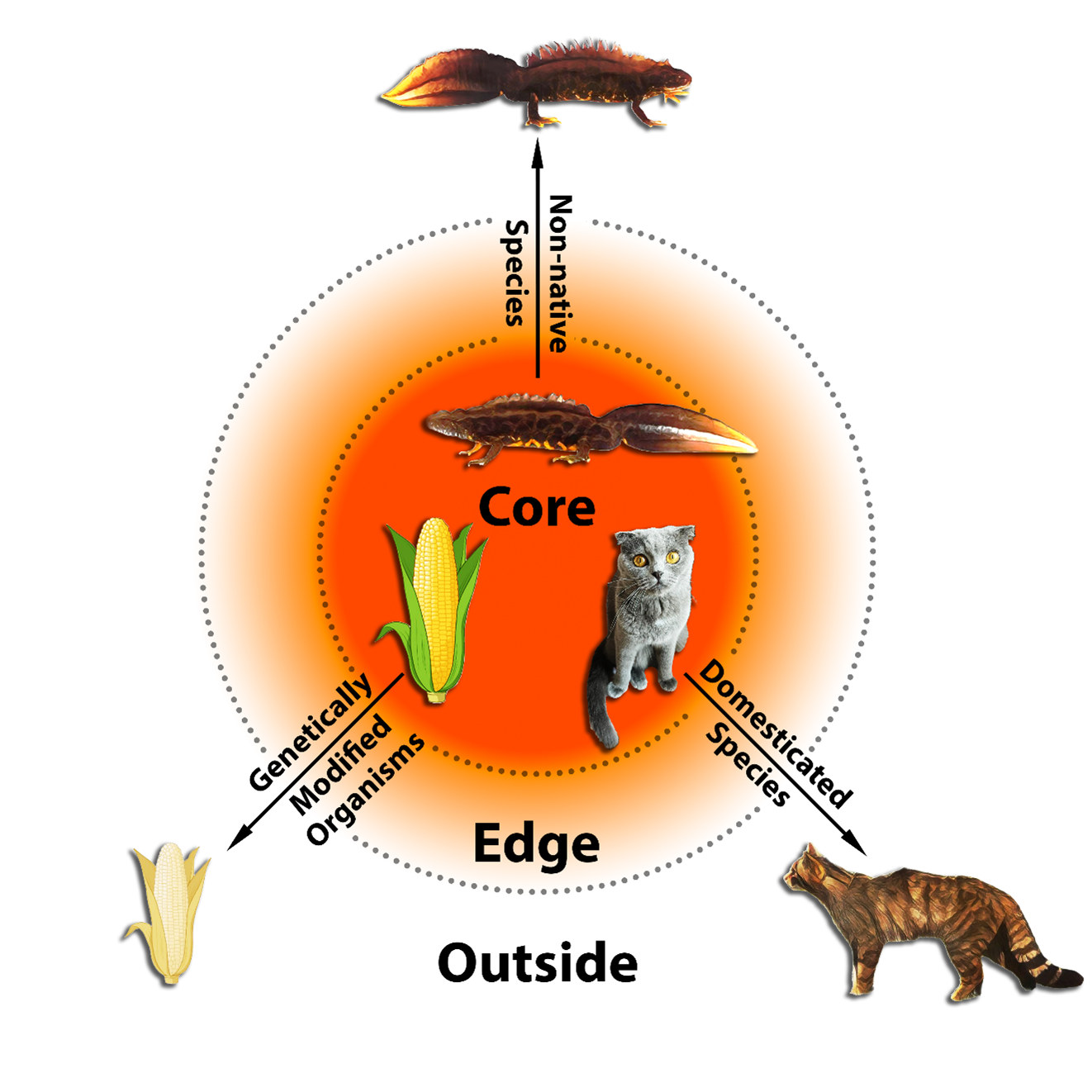
The balanced lethal system in Triturus newts is well-suited to teach biology students basic principles about natural selection and genetic linkage. This was the motivation behind a paper led by my PhD student Willem Meilink published in Ecology and Evolution. We leverage hybrids between two different crested newt species because, due to the genetic divergence between them, we can distinguish the gene versions inherited from the father and from the mother. We trace the inheritance of 30 genetic markers from throughout the genome, including multiple genes positioned on chromosomes 1A and 1B, across three generations. Our experiment perfectly illustrates that we are dealing with two tightly linked subsets of genes, one on chromosome 1A and another on chromosome 1B, that are inherited within an otherwise regularly recombining genome.

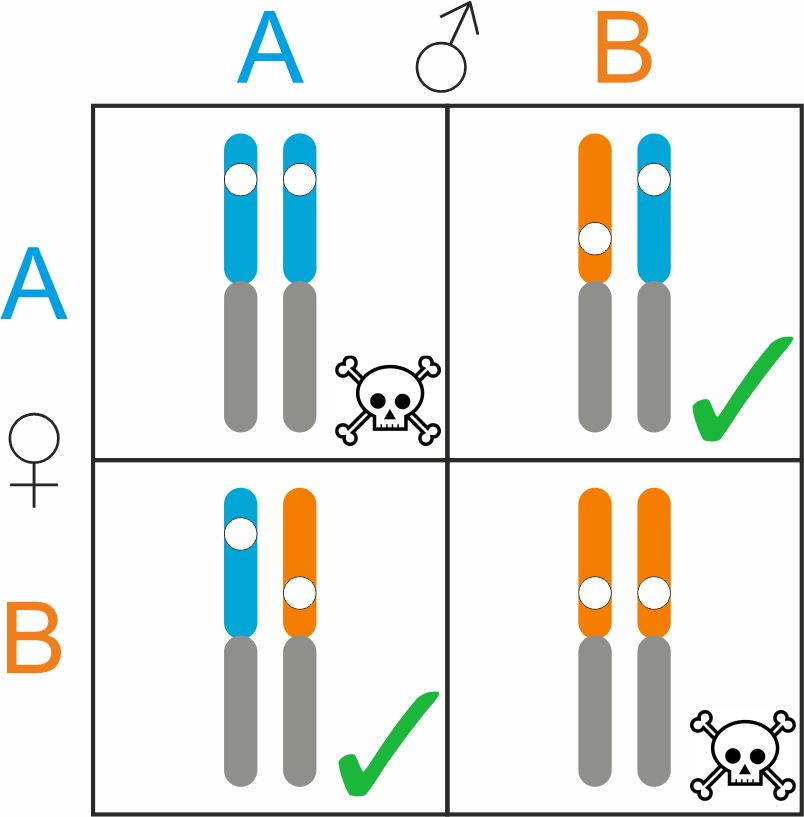
We provide evidence to support four hypotheses: 1) upon fertilization, four different genotypes are present, two healthy (1A1B = 1B1A) and two diseased (1A1A and 1B1B) ones; 2) diseased embryos that experience arrested development possess two copies of either chromosome 1A or 1B (1A1A and 1B1B); 3) healthy embryos that survive the balanced lethal system possess a single copy of chromosome 1A and a single copy of chromosome 1B (1A1B = 1B1A); and 4) genes positioned on chromosome 1A or on chromosome 1B are always inherited together (crossing over does not break up these two subsets of genes). We have been running a dedicated lab practical on this ‘Mendelian inheritance with a deadly twist’ in the second year Bachelor course Evolutionary Biology 2 at Leiden University since 2023.

Reference: Meilink, W.R.M., Cvijanović, M., de Visser, M.C., France, J., Ivanović, A., Theodoropoulos, A., Vučić, T., Wielstra, B. (2025). Exposing selection and genetic linkage in the evolutionary enigmatic balanced lethal system in Triturus newts. Ecology and Evolution 15(6): e71591.