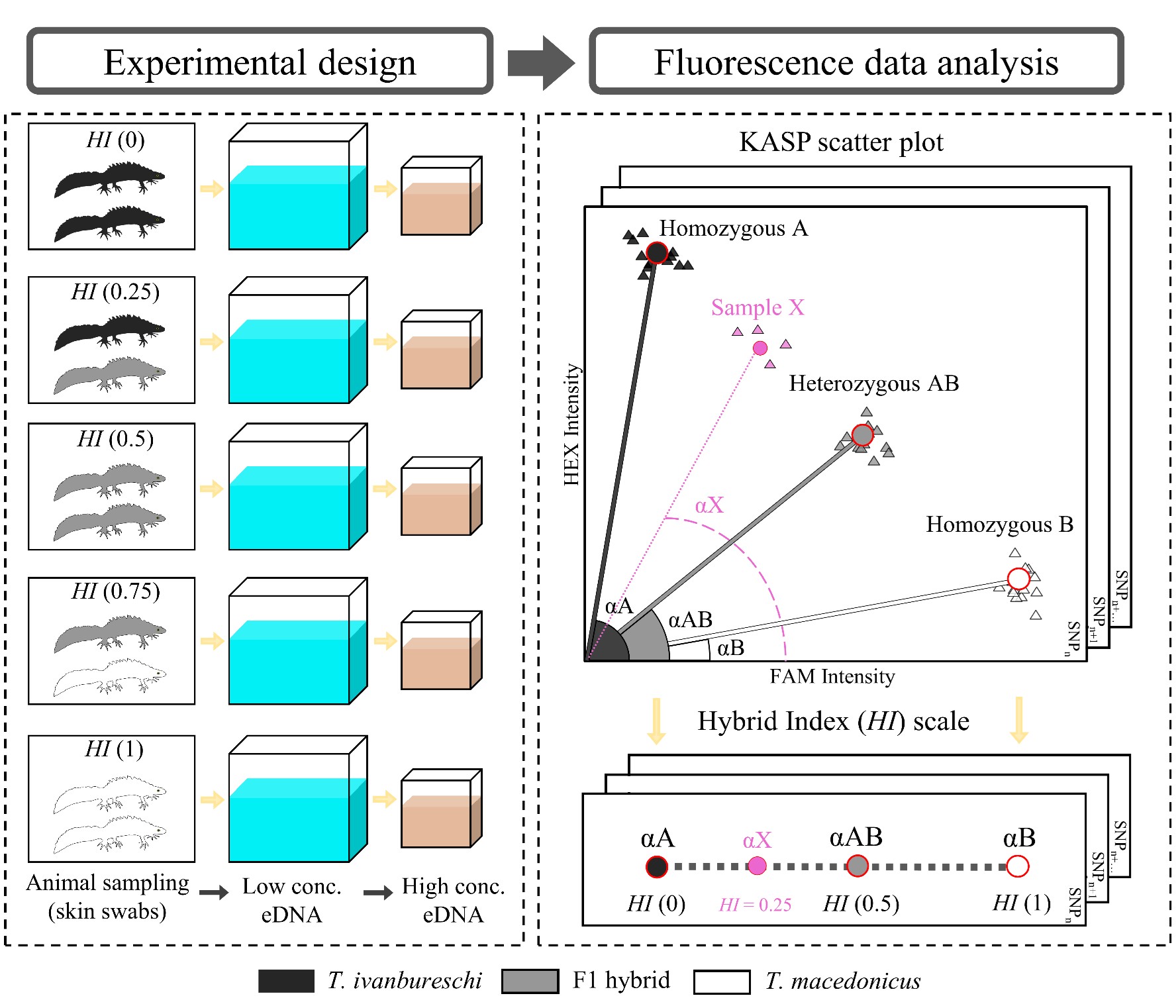
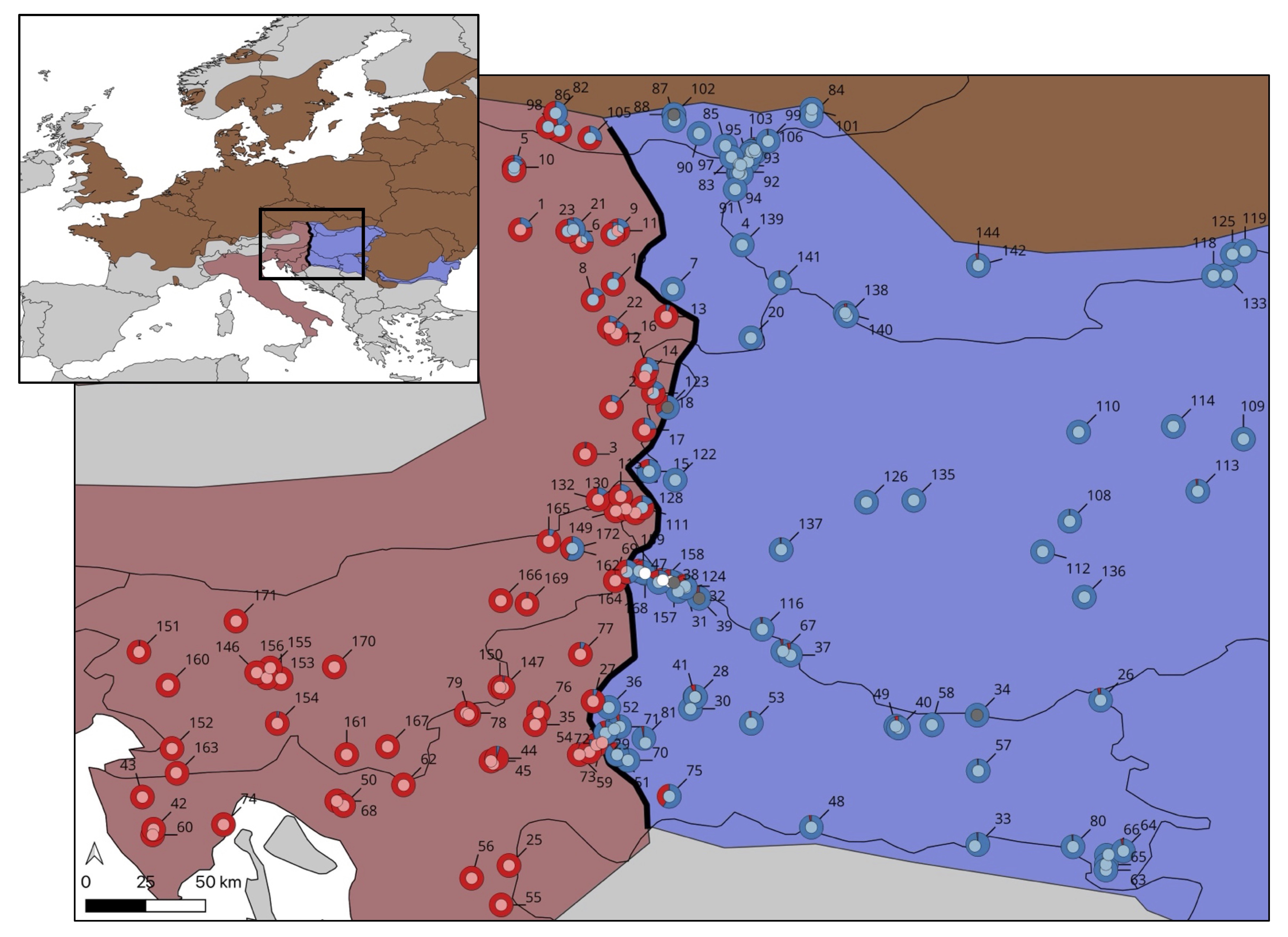
In nature, marbled and crested newts meet in France, where they manage to hybridize, despite being only distantly related. The resulting hybrid offspring are an evolutionary dead end: their fitness is practically zero. In Catalonia people have introduced the Anatolian crested newt inside the marbled newt range. Therefore, in addition to the typical threats associated with invasive species, such as competition and the spread of disease, anthropogenic hybridization should be taken into account as a conservation concern. In a paper in Global Ecology and Conservation, led by my PhD student Anagnostis Theodoropoulos, we determine genetic pollution risk in these newts.
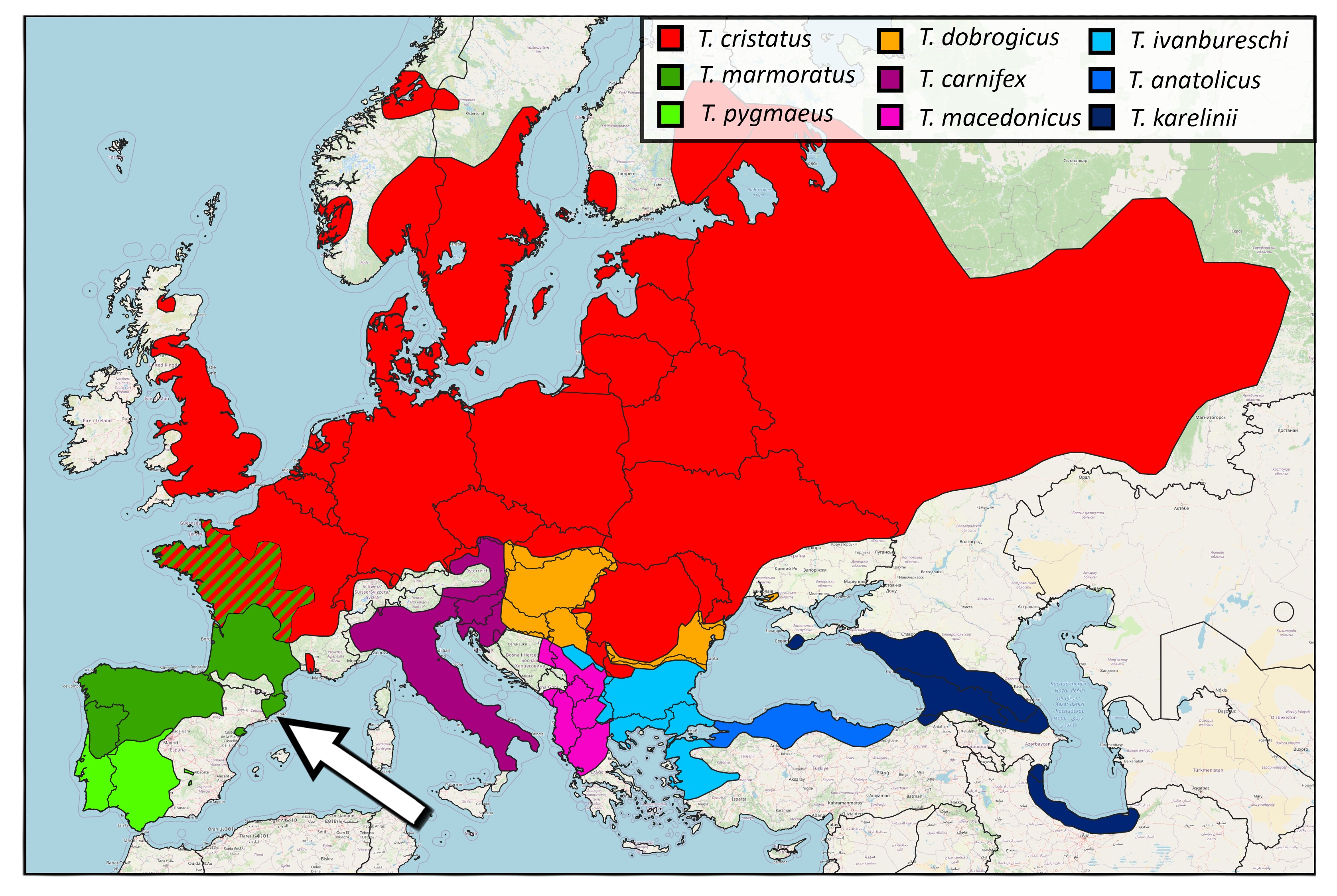
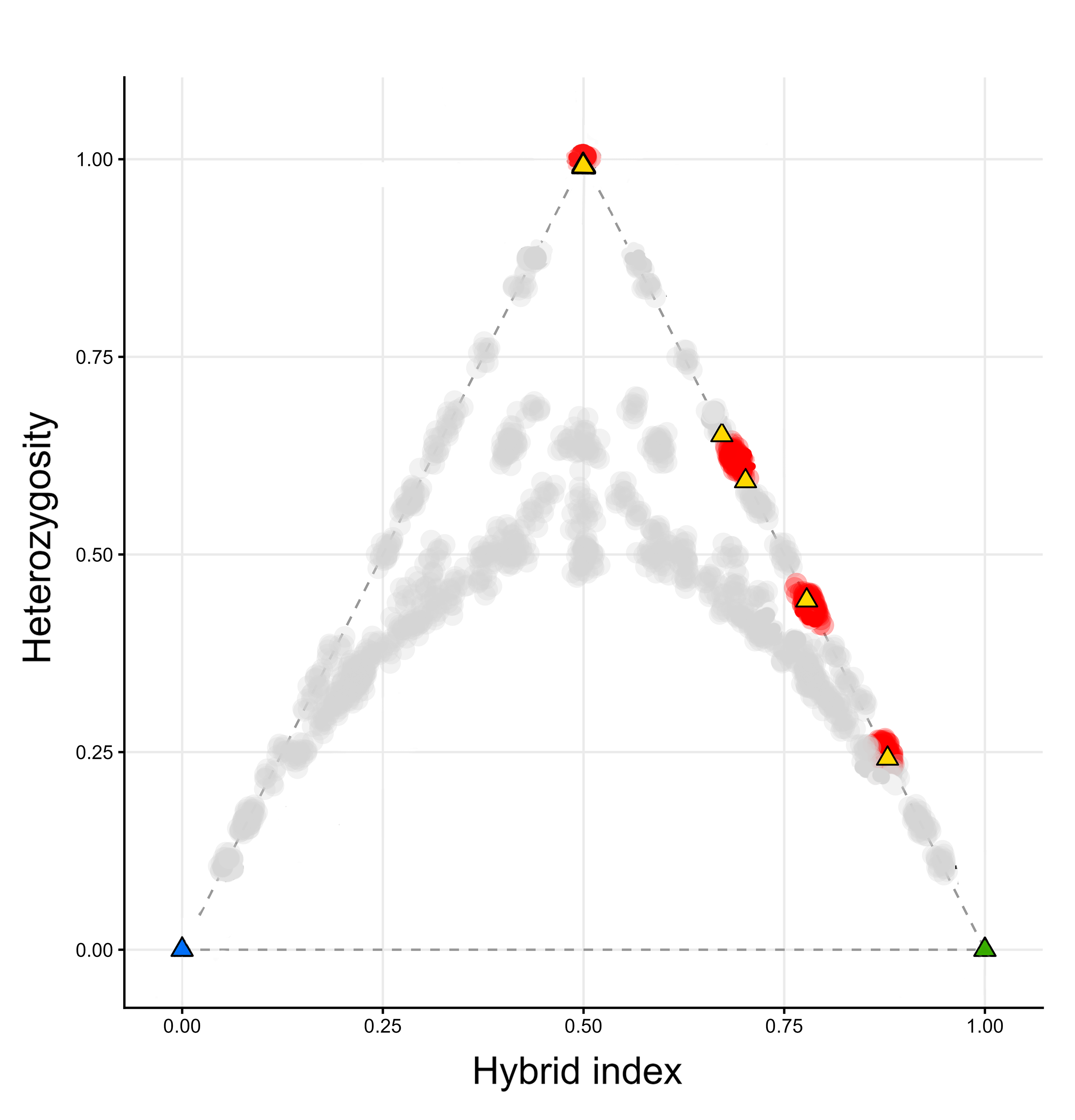
Compared to the natural hybrid zone in France, the frequency of hybridization in the human-made hybrid zone in Catalonia is considerably higher. Furthermore, the fitness of the resulting hybrids appears to be much higher than in the natural hybrid zone, because we observe a surprisingly high number of later-generation hybrids. Worryingly, all later-generation hybrids result from backcrossing towards the native marbled newt. This provides the conditions for gene flow from the invasive into the native species and we conclude that genetic pollution is a tangible risk in the Catalonian newt case. Fortunately, we caught this early.
Reference: Theodoropoulos, A., Avcı, A., Fernández-Guiberteau, D., Ferran, A., Olgun, K., Üzüm, N., Carranza, S., Wielstra, B. (2026). Genetic pollution risk in newts at the far end of the speciation continuum. Global Ecology and Conservation 69: e04308.