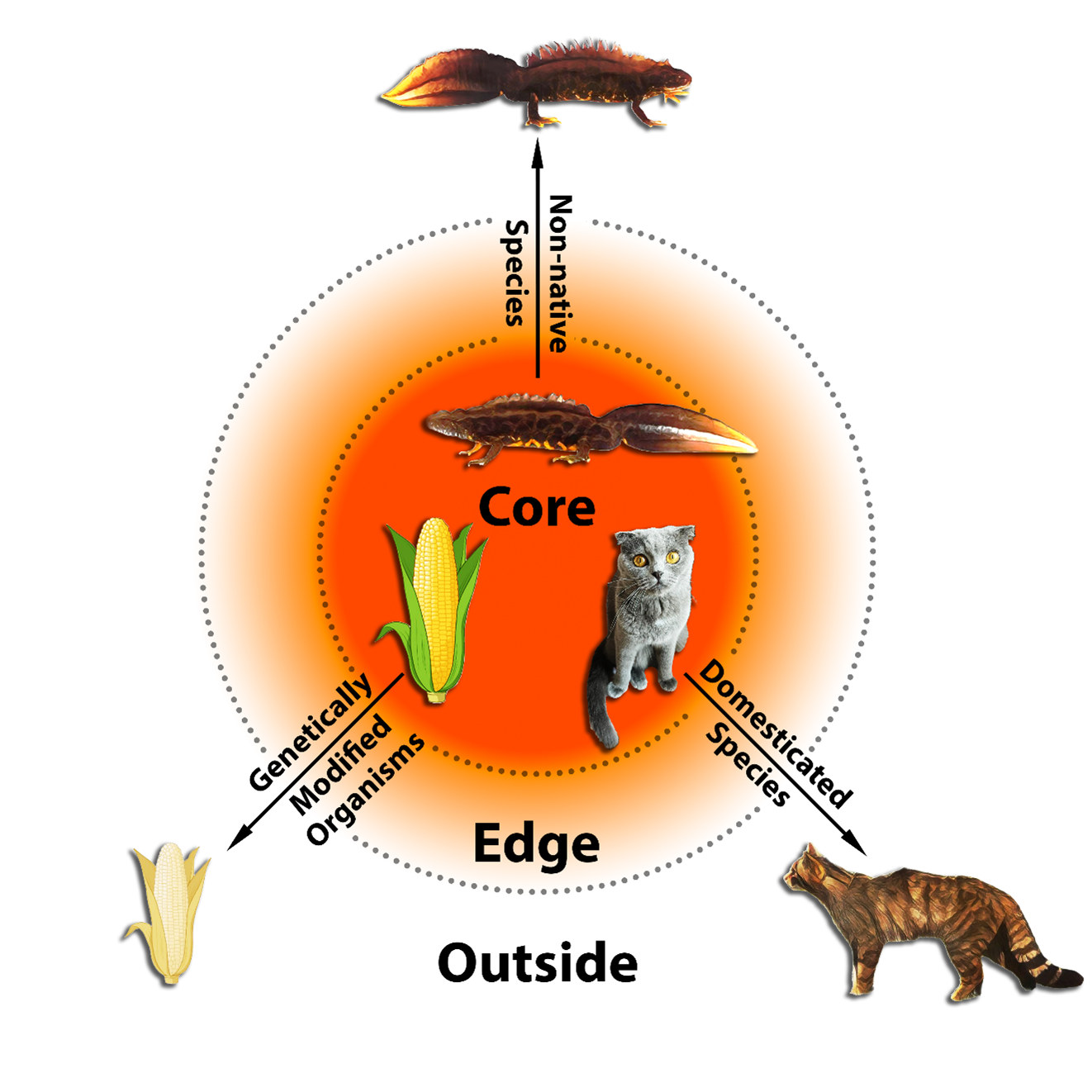
Genetic pollution, gene flow from invasive to native species species, poses a complicated conservation question. It is well-known in crested newts. Why should we care about genetic pollution? To put it bluntly, genetic pollution equals extinction. In a ‘scientists’ warning’ paper published in Discover Conservation, led by my PhD student Anagnostis Theodoropoulos, we introduce genetic pollution and stress that it is an insidious but pervasive problem. While we now have the tools available to clean up genetic pollution, policy on the problem is seriously lacking. We hope our paper serves as a call to action.
Genetic pollution at work: the orange color indicates the region dominated by non-native genetic material, whereas the white area signifies the gene pool of a native species. The gradient in between reflects non-native genes seeping into the native species.
Reference: Theodoropoulos, A., Stewart, K.A., Wielstra, B. (2025). Scientists’ warning on genetic pollution. Discover Conservation 2: 20.
This project has received funding from the Dutch Ministry of Education, Culture and Science (incentive grant), the Dutch Research Council (NWO Vidi grant VI.Vidi.213.088) and the European Union’s Horizon 2020 research and innovation programme (Marie Skłodowska-Curie grant agreement No. 655487).
The two PhD students on my ERC StG project, Manon de Visser and James France, graduated this spring. All their chapters are currently in the submission / revision process, more on the exciting science soon! For now: congratulations again!
Dr. France + committee and friends
This project has received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (Grant Agreement No. 802759).
Lissotriton vulgaris schmidtleri by Michael Fahrbach
Relationships within the newt genus Lissotriton, which includes the smooth newt, have always been difficult to interpret. Similarly, the taxonomy is quite confused: it is unclear how many species there even are. In a paper led by my former MSc students Julia Mars and Stephanie Koster we use the Triturus sequence capture protocol to bring a bit of order to this chaos. We include, for the first time, all the taxa – at least the ones that we currently know of – in a single study, just out in Molecular Phylogenetics and Evolution. We show that the palmate and not the Italian newt is sister to the smooth newt species complex. We get a lot of clarity on that smooth newt species complex and we also show where gene flow distorts relationships. We confirm the species status of some smooth newt taxa and recommend others are rather treated as subspecies. The Bosca’s newt had better not be split into two species. All these insights were made possible by the Triturus sequence capture protocol!
Reference: Mars, J., Koster, S., Babik, W., France, J., Kalaentzis, K., Kazilas, C., Martínez-Solano, I., de Visser, M.C., Wielstra, B. (2025). Phylogenomics yields new systematic and taxonomical insights for Lissotriton newts, a genus with a strong legacy of introgressive hybridization. Molecular Phylogenetics and Evolution 204: 108282.
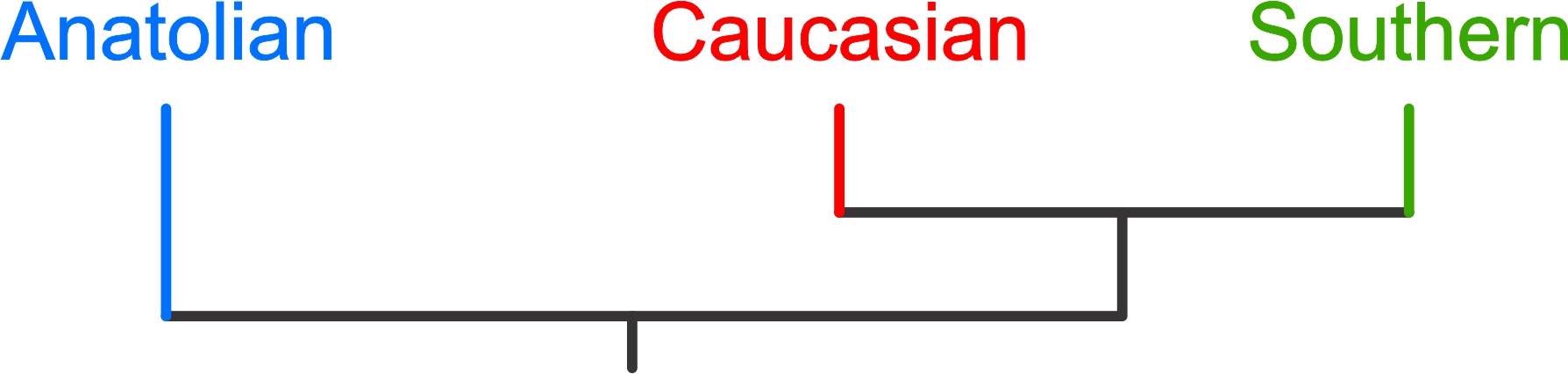
The awesome banded newt genus comprises three cryptic species that are really old and have little to no genetic exchange between them. However, the ancestral banded newt appears to have radiated into three species in a relatively short time span. Under such conditions, it is notoriously difficult to retrace the order in which species split. We certainly tried before – and failed. But now, in a paper led by my former MSc students Konstantinos Kalaentzis and Stephanie Koster and out in Molecular Phylogenetics and Evolution, we finally manage to resolve the banded newt phylogeny. The Anatolian banded newt, (currently) in contact with the Caucasian banded newt, is the first offshoot, whereas the Caucasian banded newt is the sister species of the (at the moment) geographically isolated Southern banded newt. Resolving the banded newt phylogeny was only possible because of the Triturus sequence capture protocol, which allows us to get genome-scale data for any newt!
Reference: Kalaentzis, K., Koster, S., Arntzen, J.W., Bogaerts, S., France, J., Franzen, M., Kazilas, C., Litvinchuk, S.N., Olgun, K., de Visser, M.C., Wielstra, B. (2025). Phylogenomics resolves the puzzling phylogeny of banded newts (genus Ommatotriton).Molecular Phylogenetics and Evolution 203: 108237.
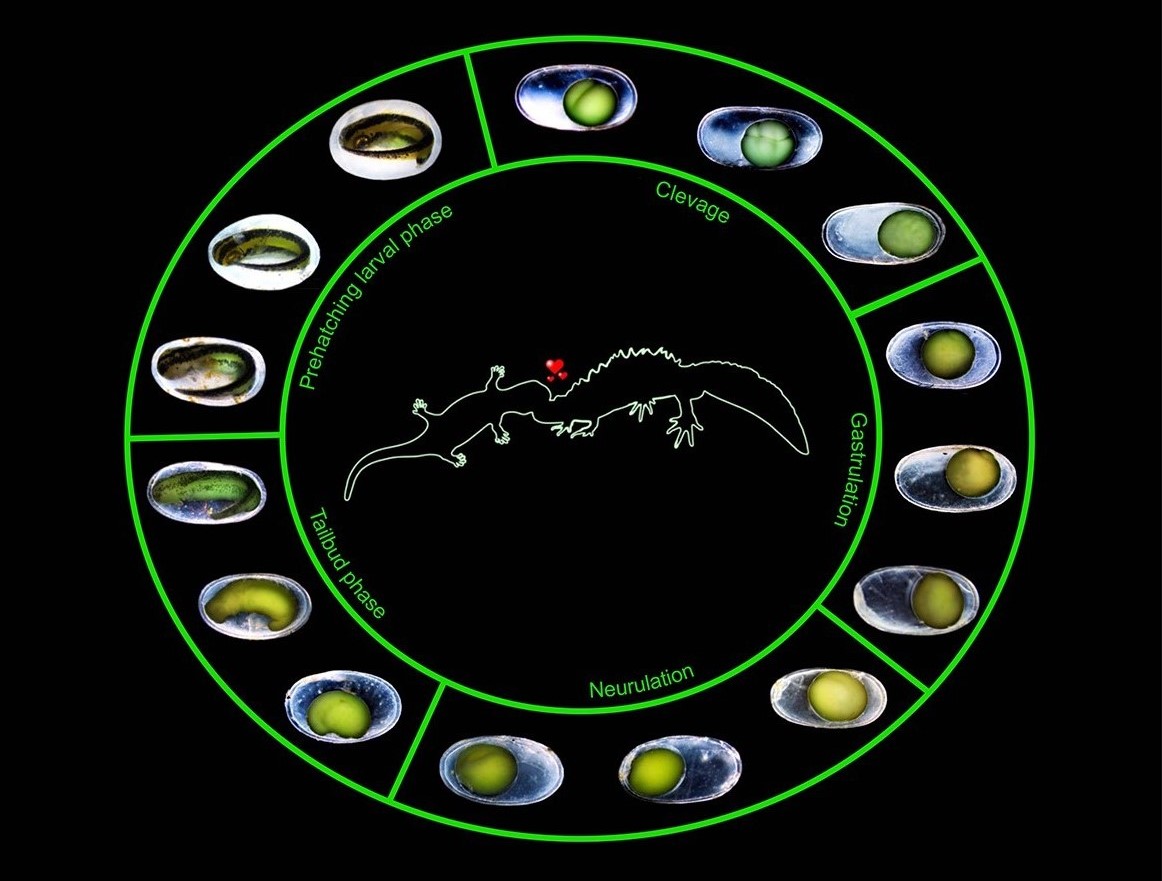
Understanding how crested and marbled newt embryos develop inside the egg is critical for my research. During this period, body shape is determined, hybrid unfitness would be expected to reveal itself, and half of the embryos are killed by a balanced lethal system! In a paper published in Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, led by my postdoc Tijana Vučić, we present a ‘staging table’ for crested newt embryonic development. A staging table is basically a summary of embryonic development, in which this continuous process is partitioned into a certain number of stages, based on outside appearance. We focus on a single crested newt species: the Balkan crested newt, Triturus ivanbureschi. Our new crested newt staging table serves as a baseline against which we can compare the embryonic development of individuals that belong to another Triturus species, that are hybrid, or that express arrested development due to the balanced lethal system. Take a look, the pictures and movies are gorgeous! You can download a high resolution PDF version of the poster below here.
This work was supported by the Serbian Ministry of Science, Technological Development and Innovation (grants nos. 451-03-47/2023-01/200007, 451-03-47/2023-01/200178, 451-03-847/2021-14/2830), the Dutch Research Council – NWO (ENW-M1 grant OCENW.M20.090), and the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant agreement no. [802759]).
In my lab we are using mtDNA barcoding to try to determine from where within the natural range introduced amphibian populations derive from. In the palmate newt, there is basically no genetic variation north of the Pyrenees Mountains. This also means that when people move newts around within this genetically depleted area, you cannot infer that from mtDNA barcoding. This is what we see in the Netherlands: the introduced populations we scanned are genetically indistinguishable from the native populations. However, if people move newts from the Iberian Peninsula to western Europe this immediately stands out. We see this in Belgium, where we detected an Iberian mtDNA barcode inside the native palmate newt range. For more details, check the paper in Amphibia-Reptilia.
Reference: Elfering, R., Mannix, S., Allain, S., Ambu, J. Crochet, P.-A., van Doorn, L., Dufresnes, C., Jehle, R., Julian, A., Kirkpatrick Baird, F., ‘O Brien, D., Secondi, J., Speybroeck, J., Theodoropoulos, A., Stark, T., Wielstra, B. (2024). Extreme genetic depletion upon postglacial colonization hampers determining the provenance of introduced palmate newt populations. Amphibia-Reptilia 45(3): 349-356.
A male pygmy marbled newt (left) and a male marbled newt. Pictures by Michael Fahrbach.
Although my work is heavily focused on crested newts, there are two marbled newt species in the genus Triturus as well. In a new paper out in Molecular Phylogenetics and Evolution, led by my former MSc student Christos Kazilas, we present a ‘next-generation phylogeography’ of marbled newts. There is remarkable little gene flow between the two marbled newt species compared to crested newts. This finding strongly supports the species status of the two marbled newts – in case anyone was still in doubt. Furthermore, both marbled newt species are composed of two genetically distinct groups. We could expose these intricate patterns thanks to the massive amount of DNA data that is easily generated with the Triturus sequence capture protocol.
Reference: Kazilas, C., Dufresnes, C., France, J., Martínez-Solano, Í., Kalaentzis, K., de Visser, M.C., Arntzen, J.W., Wielstra, B. (2024). Spatial genetic structure in European marbled newts revealed with target enrichment by sequence capture. Molecular Phylogenetics and Evolution 194: 108043.
In an article aimed at high school students for the journal Frontiers for Young Minds my former student Nienke Prins and I explain hybrid zone movement. Obviously we also mention the best example of hybrid zone movement there is: crested newts. Please have a look here.
(You can also see a previous Frontiers for Young Minds piece from our lab on balanced lethal systems here.)
This project has received funding from the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No. 655487.
In the Dune area of Meijendel, close to Leiden, a population of crested newts occurs. The history of this population is dodgy. While genetic data could not confirm that this population is introduced, the isolated position and apparent rapid expansion speak in favor of such a scenario. Previously we reported on an aberrantly colored newt from this population, with a pattern reminding of a Koi carp. Last spring we caught another odd individual, this time almost completely pale yellow.
Two pictures of either side of the same crested newt. Pictures by Anagnostis Theodoropoulos
Catching two of these weird newts in rapid succession in the same area: surely that cannot be a coincidence? I suspect this high freak-frequency fits with an introduction history for the Meijendel crested newts. The establishment of an introduced population involves similar genetic bottlenecking and inbreeding dynamics as the establishment of island populations and island populations also tend to show a higher incidence of color deviations.
Reference: Elfering, R., Bijlsma, L., Mannix, S., Plomp, S., Theodoropoulos, A., Wielstra, B. (2023). Kamsalamanders met kleurafwijking uit Meijendel. Holland’s Duinen 83: 38-39.
Three banded newt species are currently recognized. However, this was not always the case. Previously, my lab has shown that these morphologically similar species are genetically super distinct. That is why we suggested their treatment as distinct species – despite their morphological similarity. The term ‘cryptic species’ is regularly used in such a case.
Left an Anatolian (Ommatotriton nesterovi) and right a Caucausian banded newt (O. ophryticus). Pictures by Michael Fahrbach.
But is being genetically distinct enough to qualify as a species? If cryptic species meet in nature, it is possible to perform the ultimate test of species status: hybrid zone analysis. Selection against hybrids allows species to persist. Hybrid zone analysis can tell you if natural selection favors purebreds of either species over their genetically mixed offspring.
Panel a) shows the ranges of the three banded newt species. Panel b) shows all populations sampled in this study. Panel c) shows the area where the two focal species meet. Pies represent the genetic makeup of populations. There is little genetic material of one species present in populations of the other species (pies are mainly one color). Furthermore, the transition between the two species is extremely sharp; the distance between localities 20 (mainly blue) and 21 (mainly red) is only about 4 kilometers.
In a new paper published in Ecology and Evolution, we show that two of the banded newt species meet at a hybrid zone in northern Türkiye. At this hybrid zone, selection against hybrids is evidently very strong. As a consequence, the hybrid zone is extremely narrow. Our study shows that the two banded newts truly are proper species. We have already shown that, with the benefit of hindsight, it is actually possible to distinguish them based on morphological characteristics.
Reference: Kalaentzis, K., Arntzen, J.W. Avcı, A., van den Berg, V., Beukema, W. France, J., Olgun, K., van Riemsdijk, I., Üzüm, N., de Visser, M.C., Wielstra, B. (2023). Hybrid zone analysis confirms cryptic species of banded newt and does not support competitive displacement since secondary contact. Ecology and Evolution 13(9): e10442.
This project has received funding from the European Union’s Horizon 2020 research and innovation programme (under the Marie Skłodowska-Curie grant agreement No. 655487) and the ‘Nederlandse organisatie voor Wetenschappelijk Onderzoek’ (NWO Open Programme 824.14.014)